
- Technical Flexibility and Rigidity Webinar (private)
Week 8 – How dynamic environments shape cognitive flexibility
This early draft was authored by Sarah M. Pope.
Flexible problem solving has long been revered as a hallmark of human ingenuity. Yet, humans’ predilection for ‘sticking to what you know’ is evident in a wide range of behaviors and biases. That said, our global expansion and the sheer diversity of contemporaneous human cultures is a testament to our ability to flexibly adapt existing techniques or seek alternatives. The question is not ‘are humans flexible or inflexible problem-solvers?’ but rather ‘when do the benefits of being flexible exceed the costs?’ Here I propose the constrained flexibility framework for understanding how environmental variability, predictability, and harshness work in tandem to shape flexible versus inflexible strategy-use. I also distinguish between strategy switching that occurs as a result of failure and strategy switching that includes searching for and adopting a better alternative, here referred to as responsive and exploratory flexibility, respectively. Efforts to understand human cognitive flexibility have almost exclusively focused on how well we switch when we have to, rather than when, and under what contexts, we choose to. Understanding the conditions in which changing strategies is most beneficial may help explain how our species strikes the balance between maintaining working strategies and finding or creating better ones.
Introduction
I recently heard a Congolese fable describing how the antelope, a weak but wily animal, became the king of the jungle. It had been decreed that whichever animal was strong enough to shoot an arrow through the trunk of the largest tree would win the throne. The elephant and the leopard, the strongest of the animals, practiced and practiced. But on the day of the competition their arrows barely pierced its bark. They laughed when the antelope stepped up to take his turn. However, knowing that he would never be strong enough, the antelope had come up with a different strategy. In the days before the event, the antelope convinced his friend, the bee, to drill a hole through the tree in a place that only he would know. On the day of the competition, the antelope took aim and shot the arrow through the hole, thereby winning the throne (told by Francy Ntamboudila, 2020). The leopard and the elephant failed because they relied solely on their might, whereas the antelope’s flexible thinking showed his true strength.
Flexible behavior is an adaptive response to dynamic environments. As challenges arise or opportunities shift, strategies must be updated in order to meet changing demands. The umbrella term behavioral (or phenotypic) plasticity refers to an individual’s ability to produce different behaviors in response to environmental changes (Fox & Weber, 2002; Snell-Rood, 2013). This chapter will focus on the cognitive mechanisms underlying flexible behavior, or cognitive flexibility. Cognitive flexibility is the ability to adaptively select between known solutions and innovated or acquired novel solutions in response to relevant environmental changes (Laureiro-Martínez & Brusoni, 2018; Pope, 2018; Ueltzhöffer et al., 2015). Thus, cognitive flexibility is defined as the – contextually mediated – optimal balance between repeating or returning to a familiar strategy and searching for an alternative, with suboptimal strategy-selection occurring as a result of either inflexibility, an inability to switch from an active strategy to a better one, or overflexibility, an inability to maintain the best strategy.
Environments fluctuate predictably, with daily or seasonal cycles, and unpredictably, in response to stochastic events like fires or floods. An animal’s lived environment is also a product of its habitat, which may span multiple, even micro-, climates, and its ranging patterns, which might regularly extend into novel territory. Flexible behavior is especially important for long-lived, mobile species, like humans, who must cope with changing environments throughout their lifetimes (Dingemanse & Wolf, 2010; Sol et al., 2016; van Schaik, 2013; Vicente & Wang, 1998a). Indeed, humans exhibit an unprecedented degree of behavioral diversity (Fogarty et al., 2015; Hill et al., 2009) which allows us – for better or for worse – to inhabit and modify over half of earth’s landmass (Henrich & McElreath, 2003; Vitousek et al., 1997), suggesting that, at least as a species, humans are quite capable of updating their existing strategies to meet environmental needs. However, on an individual level, flexibility is not always evident.
Like the elephant and the leopard, humans are often blinded by familiar strategies or known solutions. In a classic example, children and adults learned to solve a set of arithmetic ‘water jar’ problems using a somewhat tedious, four-step solution. After they solved six examples in this way, they were given four problems that could be solved either by the learned solution or by a more efficient, one-step alternative (Luchins, 1942). Luchins tested thousands of American adults and children, using various manipulations, and consistently found that a large majority of participants continued to use their learned strategy despite the presence of a better alternative (Luchins, 1942; Luchins & Luchins, 1950). Termed ‘cognitive set,’[1] the propensity for familiar strategies to occlude - even more efficient – alternatives, has been demonstrated across a wide variety of contexts including strategic reasoning (Bilalić et al., 2008), design and engineering (Chrysikou & Weisberg, 2005; Jansson & Smith, 1991), mathematics (Crooks & McNeil, 2009; Lemaire & Leclère, 2014; Wertheimer, 1945), sequential problem solving (Jacobson & Hopper, 2019a; Pope et al., 2015, 2019, 2020; Watzek et al., 2019), insight problem solving (Hanus et al., 2011; Öllinger et al., 2008), and functional fixedness paradigms (Adamson, 1952; Duncker & Lees, 1945; German & Barrett, 2005). We are faced with a contradiction, on one hand humans are profoundly adaptive, the inventors and modifiers behind a technological revolution, but on the other hand, we are dismally conservative, either unwilling or unable to move beyond the tried and true. How can humans be simultaneously flexible and inflexible? Here, I suggest that the answer lies in a different question, rather ‘when do the benefits of being flexible exceed the costs?’
In this chapter, I begin by discussing the relative risks and benefits of repeating a current strategy (staying), switching to a known alternative (switching), or searching for a novel alternative (searching), highlighting that the optimal balance between the three is highly dependent on exogenous demands. Next, building off of existing theories of human cognitive evolution, I propose a framework for how environmental variability, predictability, and harshness might work in concert with one another to shape cognitive flexibility.
Costs and benefits of staying, switching, and searching
Why change strategies?
Changing strategies is beneficial when an alternative strategy is better than the current strategy. Under forced-switchconditions, when the current strategy no longer works, a shift in goal or approach is clearly adaptive as the value of the current strategy has become zero. Switching or searching that occurs under forced-switch conditions will be henceforth referred to as responsive flexibility. Strategies can become temporarily or even permanently ineffective for a number of reasons. Capabilities, goals, and opportunities shift throughout development and in response to exogenous changes in the environment like predation or other mortality risks, seasonal or spatial fluctuations in weather and resource availability, as well as downstream effects of climatic variability or anthropogenic disturbance. When a current strategy stops working, adaptive behavior might entail the discovery or innovationof a new technique, or the application of another known strategy to a new context. Under forced-switchconditions, inflexibility always results in failure. The majority of human cognitive flexibility research makes use of forced-switch metrics, in the form of intra- and extra-dimensional set-shifting (Doebel & Zelazo, 2015; Zelazo, 2004) or task-switching paradigms (Meiran, 2010; Monsell, 2003; Sakai, 2008). Responsive flexibility, or rather inflexibility, is then measured by the extent to which actors perseverate with their current strategy after it stops working. Changing strategies under forced-switch conditions is considered an integral part of executive functioning (Doebel & Zelazo, 2015; Miyake et al., 2000).
The other context in which flexible behavior can be beneficial is when a current strategy is less efficient than an alternative. Under optional-switch conditions, rather than avoiding failure, changing strategies can reduce inefficiency but not always. Switching or searching that occurs under optional-switch conditions will be henceforth referred to as exploratory flexibility. Exploratory flexibility offers an important mechanism for understanding human ingenuity and cumulative culture, which necessarily builds upon existing techniques. However, within psychology, flexibility under optional-switch conditions is rarely measured. Existing metrics include the Water Jar (described above) and other learned-sequence tasks (Luchins, 1942; Luchins & Luchins, 1950; Pope et al., 2020; Watzek et al., 2019) along with some task shifting (Ardiale & Lemaire, 2012; Arrington & Logan, 2004; Lemaire & Leclère, 2014), token exchange (Hopper et al., 2011; Van Leeuwen et al., 2013; van Leeuwen & Call, 2017), and extractive foraging tasks (Davis et al., 2019; Jacobson & Hopper, 2019b; Price et al., 2009). However, the balance between exploiting useful strategies and exploring alternatives, has been extensively studied within foraging (Charnov, 1976; Cohen et al., 2007; Stephens & Krebs, 1986) and decision making research (Acuna & Schrater, 2007; Fischhoff & Broomell, 2020; Gittins & Jones, 1979; Payne et al., 1994; Peterson & Verstynen, 2019), wherein it is measured using a range of reinforcement learning and sequential decision-making paradigms, like patch-leaving and multi-armed bandit tasks (see Averbeck, 2015 for a review). In optional switch conditions, the benefit of selecting a better strategy is pitted against the computation and search effort required to compare alternatives, combined with the opportunity costs incurred by learning or switching delays.
Why repeat a strategy?
Failure to switch strategies occurs in many contexts, and is not always maladaptive. In forced-switch conditions, perseveration occurs when a previously successful strategy continues to be evoked, despite evidence that it no longer works (Floresco, 2011; Schillemans, 2011). Perseveration can be adaptive if the failure is transient, or the causal mechanisms are unclear (i.e. the failure might have been a fluke). In optional-switch conditions, where multiple strategies are available, changing strategies is only valuable if it leads to adopting a better one. In other words, it would be maladaptive to switch if the chosen alternative was not more efficient than the current strategy – or if the efficiency benefit of the chosen alternative failed to exceed the time invested in finding, selecting, and honing it. Another benefit of inflexibility is that it can help maintain or refine useful strategies, reallocating the effort that might be spent searching towards skill practice and eventually specialization (Dingemanse & Wolf, 2013). Additionally, learning and maintaining a new strategy is neurologically expensive (Laughlin & Sejnowski, 2003). It seems likely that repeating a strategy is an adaptive default-approach, but one that should be deviated from when the benefits of finding an alternative exceed the costs (Duckworth, 2010).
Dealing with ambiguity
When all possible strategies are known, optimal behavior is simply a matter of switching to or maintaining the best one. However, we do not occupy nor have we evolved within unambiguous environments. Under conditions of uncertainty, the relative risks and rewards of potential strategies are often unclear. To make an optimal decision, one could search for all possible strategies and test their relative risks and payouts, but the time and effort spent gathering these data may quickly outweigh the benefit of selecting the better alternative, at least in the short-term. Heuristics, such as ‘always choose the second-cheapest wine’ (McFadden et al., 1999), are individuals’ set of guiding principles which support decision-making under uncertainty, often by systematically ignoring some of the available information (Gigerenzer et al., 2011). The computational requirements of perfect decision-making are immense and heuristics can be useful cognitive shortcuts, but they lead to predictable biases and errors (Kahneman et al., 1982; Tversky & Kahneman, 1974), especially when cognitive capacity is limited (Cash-Padgett & Hayden, 2019). Behavior guided by heuristics is not often optimal, but on average, it should reach an acceptable level of efficiency, at least within the environment in which it was formed (Fawcett et al., 2013).When the relative risks and benefits of all possible strategies is unknown or unclear, the decision to repeat a strategy, switch to a known alternative, or search for a new one is likely guided by heuristics, which are, ideally, tuned to optimize decision-making in that environment (Todd & Gigerenzer, 2007; Vicente & Wang, 1998b). To understand cognitive flexibility we must first consider how exogenous pressures, like variability, predictability, and harshness might serve to shape our biases towards staying, switching, and searching.
The constrained flexibility framework
When considering how environmental forces shape cognitive flexibility, we can make use of existing hypotheses which posit that adaptive cognition is heavily influenced by environmental variability, the extent to which the environment changes over time or space, predictability, the temporal regularity of changes or the degree to which they are correlated, and harshness, the level of consequence elicited by strategy failure (see Box 1). These hypotheses offer important perspectives; however, they often either conflate variability, predictability and harshness, or consider their effects in isolation.
Here, I propose that recognizing how variability, predictability, and harshness work in concert with one another to regulate cognitive flexibility is integral to predicting when strategies should be repeated or changed, and if change is required, how the relative costs and benefits of responsive vs exploratory flexibility are balanced. Recall that responsive flexibility consists of switching or searching as a result of strategy-failure and exploratory flexibility is switching or search that occurs despite already having a working strategy. Exploratory flexibility is risky but here I argue that under certain circumstances, the adaptive benefit of potentially finding or innovating a better solution is enough to offset the costs.
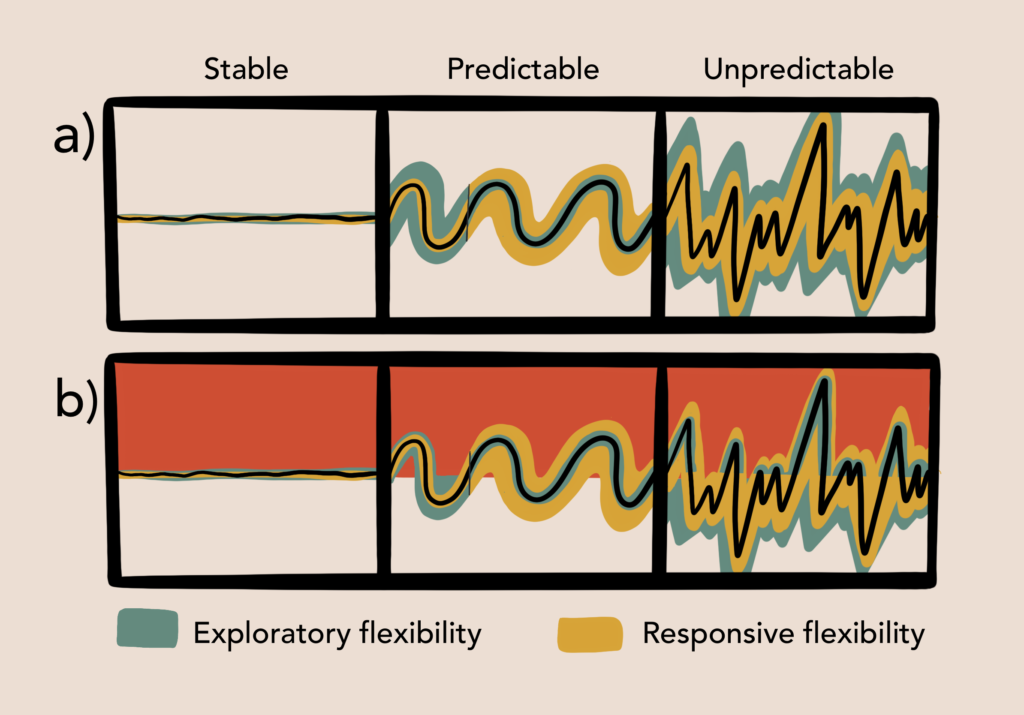
The constrained flexibility framework unifies existing hypotheses regarding the impacts of environmental variability, predictability, and harshness, on behavioral flexibility, taking into account their unique and combined effects, in order to predict the circumstances under which exploratory flexibility might contribute to enhanced cognitive flexibility. Specifically, it suggests that exploratory flexibility is suppressed in harsh, stable or predictably variable environments, but may be a valuable tool in unpredictable environments, so long as harshness is low.
Specifically, in stable environments, the usefulness of a strategy does not change over time and so responsive flexibility is not needed. Once an optimal strategy is found, there is little benefit derived from maintaining, using, or seeking alternatives. Thus, under stable conditions, exploratory flexibility is maladaptive – and should be suppressed – so long as a working strategy can be maintained. In variable environments, the usefulness of a strategy can either change regularly, as in predictably variable environments or irregularly, as in unpredictably variable environments. In variable environments, regardless of harshness, responsive flexibility is critical for handling situations where no working strategy is known (like in novel environments) or when a previously effective strategy stops working. However, the conditions which predict exploratory flexibility are less clear. In predictably variable environments, exploratory flexibility may only be useful initially, during a strategy accumulation stage. In this way, a range of strategies might be acquired and then, if memory capacity is high enough, switched between as needed. Extended explorative behavior is not useful in predictably variable environments because even though optimal strategies rotate over time, the set of optimal strategies is unchanged. Thus, cognitive resources should be devoted to maintaining and honing these working strategies rather than searching for alternatives. In unpredictably variable environments, previous strategies may or may not work well, or even at all. Here increased exploratory flexibility might serve to minimize delays in updating to new optimal strategies or in some cases, result in the discovery or innovation of a novel, more efficient technique. Importantly, both predictably and unpredictably variable environments can exist partly or wholly in states of harshness, which places a penalty on exploratory flexibility commensurate with the severity of the consequences of failure. Responsive flexibility is unaffected by harshness, because it only occurs when a previous strategy stops working and thus no amount of harshness will outweigh the benefit of switching or searching for an alternative. The following sections will describe existing hypotheses and evidence supporting the constrained flexibility framework. See Figures 1-2 for a graphic representation of these dynamics.
Box 1. Hypotheses regarding environmental impacts on adaptive cognition.
The idea that environmental challenges, such as variability, predictability and harshness, act as major selection forces in shaping cognition is certainly not new. Many researchers have proposed a role of exogenous variation, or novelty, in cognitive evolution. For example, according to the behavioral drive hypothesis (Wyles et al., 1983), expansion into novel habitats requires larger brains that are capable of handling the accompanying challenges, such as locating, identifying, and procuring unknown resources or avoiding risks that stem from novel climates, landscapes, and predators. And the adaptive flexibility hypothesis proposed that an initial period of high behavioral diversity is essential to adjusting to a novel habitat, with a subsequent decline as successful behavioral variants are honed (Wright et al., 2010).
Other hypotheses have focused on the role of encephalization on adaptive behavior. For instance, the cognitive buffer hypothesis (Allman et al., 1993; Deaner et al., 2003) and the brain size–environmental hypothesis(Sol et al., 2005) suggest that increased cognitive capacity, supported by larger brains, acts to buffer or as a means of handling changing environments. Supporting these claims, there is evidence that in primates, carnivorous mammals, and birds, neocortex size correlates with rates of innovative problem-solving (Benson-Amram et al., 2016; Lefebvre et al., 2004; Simon M. Reader & Laland, 2002; Simon M. Reader & MacDonald, 2003).
The role of harsh environments in promoting adaptive change (recently reviewed by van de Pol et al., 2017) is prominent in many well-known hypotheses aimed at understanding animal behavior and speciation, including habitat theory (Vrba, 1995), the savannah hypothesis (Dart & Salmons, 1925; deMenocal, 1995), and niche construction (Odling-Smee, 1988; also see Laland et al., 2016). For example, in the necessity is the mother of invention hypothesis (K. N. Laland & Reader, 1999; S M Reader & Laland, 2001) innovative adaptations are thought to only arise when needed. However, this stands in opposition to the spare time hypothesis which suggests that innovation occurs in times of low stress, due to lower consequences of failure (Kummer & Goodall, 1985).
Taking into account the role of environmental variation, rather than just extremes, Potts’ (1996) proposed the variability selection hypothesis, which suggests that in response to fluctations in temperature, aridity, and water abundance, early hominins developed a suite of cognitive traits that allowed them to not only move to more suitable environments but also develop flexible approaches to buffer local environmental instability (Grove, 2011; Potts, 2012). And in support of this, through a combined analysis of paleoenvironmental and archaeological records of the last 5 myr, Potts & Faith (2015) demonstrate considerable overlap between periods of high environmental variability and key behavioral adaptations and speciation events in the evolution of Homo (see also Snell-Rood, 2013).
When to stay, switch, or search
“In an uncertain and changing environment, where values of all potential options are unknown and/or the values of these options change over time, one must adapt by flexibly alternating between exploration and exploitation in order to maintain efficient performance over time and to keep track of the state of the environment.” - (Addicott et al., 2017, p 1932)
Predictable versus unpredictable environments
Both predictable and unpredictable variation are common in the natural world. Predictable variation can be seen in the many cycles which govern our climate on daily, monthly, and annual bases. Seasonal fluctuations, especially, might be drastic and require a range of behaviors throughout the year. However, seasonal transitions occur smoothly and predictably. Riotte-Lambert & Matthiopoulos (2020) suggest that when environmental predictability is high, learning and memory are advantageous, because they allow existing strategies to be recalled and activated when needed, reducing the need for strategy search and its accompanying costs (also see: Colwell, 1974; Milton, 1981). Indeed, in several decision-making tasks, where participants must choose to either exploit a current resource or explore alternatives, more exploratory behavior occurs in response to unpredictable or stochastic, compared to stable, reward structures (Behrens et al., 2007; Navarro et al., 2016; Speekenbrink & Konstantinidis, 2015).
Even within mostly predictable environments, some volatility is inevitable. It seems likely that humans’ suboptimal behavior in optional-switch paradigms, where familiar strategies occlude better alternatives, might be the result of reduced exploratory flexibility that occurs when the environment is (in these cases, inaccurately) considered to be fairly stable (Bilalić et al., 2008; Luchins, 1942). In these studies, participants under-searched their environments and this seems to occur in humans’ everyday lives as well. Following two days of public transportation strikes, an estimated 5% of London commuters, who had been forced to switch from their normal routes by station closures, did not return to their previous routes; suggesting that the need to search for an alternative strategy resulted in their finding a better one (Larcom et al., 2017).
Inaccurate stability judgements may also help explain cross-cultural differences in cognitive set. Pope et al. (2019) found that Namibian Himba participants were ~4 times more likely to find and use a more efficient ‘shortcut’ strategy than Americans on a touch screen sequential problem-solving task. It is possible that unfamiliarity with computer games led Himba participants to consider the game less predictable than their American counterparts. However, another intriguing possibility is that if decision-making heuristics are somewhat domain-general, unpredictability in other areas of Himba participants’ lives might have result in a lowered threshold for exploratory search. This possibility aligns with other research which found that younger children exhibited more exploratory flexibility than adolescents and adults (German & Defeyter, 2000; A Gopnik et al., 2015; Pope et al., 2015). Indeed, young children may be more prone to exploratory flexibility as they are in a constant state of strategy acquisition, wherein predictive decision-making, based on reasoning and executive functioning, may still be out of reach (Alison Gopnik, 2020; Ionescu, 2017).
Harsh environments
Thus, exploratory flexibility may be a valuable tool for handling environmental changes, especially those that are unpredictable, but it comes with a price. Searching for a novel strategy can take time, and may result in failure. If a viable alternative is found, there may be a learning delay before it can be enacted, or attempts to learn it could fail. Finally, the net benefit of using the alternative strategy might be lower than the previous strategy, based on the time invested in finding, learning, and using it.
In harsh environments, time is limited and the consequences of failure are high. It seems likely therefore that harshness places severe constraints on exploratory flexibility. For example, in a modified water-jar paradigm, Beilock & Decaro (2007) found that participants with higher working memory were more likely to use the suboptimal four-step solution under conditions of high stress. Similarly, Wilson et al. (2017) found that humans decrease their information seeking behavior when time horizons (the maximum number of choices or task duration) are short. Additionally, Averbeck (2015) reviews a number of patch-leaving and multi-armed bandit tasks demonstrating that search is less beneficial when time horizons are short and the consequences of failure are high. Thus, exploratory flexibility should only occur in harsh environments when it is absolutely necessary – when no other options exist.
An analysis of commercial fishing vessel positional records showed that in normal conditions, more exploration did not increase fishing performance. However, during a major disturbance, which resulted in the closure of the most-used fishing grounds, boats that had explored during normal conditions outperformed their non-exploratory peers, and were more likely to continue fishing despite the disturbance (O’Farrell et al., 2019). It follows that strategy accumulation may be most beneficial when an individual spends some, but not all of its time, in harsh environments. Addicott et al. (2017) suggested that increased exploration and strategy accrual during periods of moderation, when the benefit of a possible alternative exceed the search costs, might result in useful back-up strategies that could later be deployed during periods of harshness. They even expanded this idea to account for risky behavior during human adolescence, noting that it likely provides valuable information that can be utilized later in adulthood (Mata et al., 2013). For example, hunter-gatherer toolkits are significantly predicted by proxies for risk of resource failure, suggesting that the accumulation of more tools serves to buffer harshness (Collard et al., 2005).
Thus, in alignment with the ‘necessity is the mother of invention’ hypothesis, constrained flexibility predicts that when no other options are available exploratory flexibility should kick in, even in harsh environments. However, consistent with the spare time hypothesis, it notes that exploration during periods of relative calm can also provide fitness benefits, even at later times. Exploratory flexibility is a powerful, but costly tool. Next, we look at ways in which the price of exploration can be mitigated.
Mitigating costs of exploration
Directed versus random exploration
I’m considering a section on directed vs random exploration here.
Socially Acquired Information
In humans, the most prevalent means of reducing exploration costs is by copying successful strategies from other individuals. The Cultural Brain hypothesis suggests that larger brains are selected for their ability to acquire adaptive information via both asocial and social learning (Muthukrishna, 2015). Evidence from simulations suggest that the combination of asocial and social information can be very adaptive (Acerbi & Parisi, 2006; Muthukrishna et al., 2018). Especially in technologically advanced settings, where one cannot easily devise the underlying mechanism, social learning is an important tool (Froese & Leavens, 2014).
Interestingly, humans may be over-reliant on socially acquired strategies, as is evidenced by our proclivity for over-imitation (Hoehl et al., 2019; Horner & Whiten, 2005). Importantly, within the constrained flexibility framework, access to useful socially-acquired information would act to suppress exploration because the costs and benefits of individual exploration are often unknown, and therefore eclipsed by the low cost and known benefit of a socially acquired strategy. Indeed, Bonawitz et al. (2011) found that children who were taught how to play with a novel tool were far less exploratory than naïve peers. However, this does not seem to be maladaptive overall; novel contexts always require strategy search, but if useful social information can be efficiently acquired, then search cost can be almost entirely avoided.
[Considerations regarding reliance on social information, especially in the face of harshness, to be expanded here. Also to discuss the inherent social value, in terms of conformity and cultural cohesion, that arise from using socially-acquired strategies.]
Concluding remarks
In reality, environments exist along continua – from stable to flexible, predictable to unpredictable, mild to harsh. Humans occupy a range of ecological and cultural environments simultaneously and the concepts of variability, predictability, and harshness depend on the domain and time scale being considered. Furthermore, our exposure to environmental pressures is confounded by our behaviors, such as movement patterns or our ability to buffer variability (e.g. niche construction; see Chevin & Hoffmann, 2017 for discussion). By differentiating between strategy changes that occur in response to failure, or responsive flexibility, versus those that occur proactively, or exploratory flexibility, the constrained flexibility framework provides a theoretical basis for understanding how our species balances the need to maintain strategies that work with the benefit of finding or creating better ones. [To be expanded and tied back into the introduction more succinctly.]
Terminological question for the other authors
In this chapter I have used a common definition of cognitive flexibility, which describes it as the optimal balance between repeating or returning to a familiar strategy versus trying a novel alternative. However, this definition becomes problematic when one simplifies it into: the optimal balance between being inflexible (continuing to use a strategy) and being flexible (switching strategies). Indeed, one could be “cognitively flexible” while acting “inflexibly” if the strategy that is being repeated is the best option. I would like to ask if the other authors are aware of alternative terms that are already in use or, if not, request feedback on the use of ‘adaptive cognition’ as a replacement for ‘cognitive flexibility.’
References
Acerbi, A., & Parisi, D. (2006). Cultural transmission between and within generations. Journal of Artificial Societies and Social Simulation, 9(1), Article 1.
Acuna, D., & Schrater, P. (2007). Bayesian Modeling of Human Sequential Decision-Making on the Multi-Armed Bandit Problem. Bernoulli, 2065–2070.
Adamson, R. E. (1952). Functional fixedness as related to problem solving: A repetition of three experiments. Journal of Experimental Psychology, 44(4), 288–291.
Addicott, M. A., Pearson, J. M., Sweitzer, M. M., Barack, D. L., & Platt, M. L. (2017). A Primer on Foraging and the Explore/Exploit Trade-Off for Psychiatry Research. Neuropsychopharmacology, 42(10), 1931–1939. https://doi.org/10.1038/npp.2017.108
Allman, J., McLaughlin, T., & Hakeem, A. (1993). Brain weight and life-span in primate species. Proceedings of the National Academy of Sciences, 90(1), 118–122. https://doi.org/10.1073/pnas.90.1.118
Ardiale, E., & Lemaire, P. (2012). Within-item strategy switching: An age of comparative study in adults. Psychology and Aging, 27(4), 1138–1151. https://doi.org/10.1037/a0027772
Arrington, C. M., & Logan, G. D. (2004). The cost of voluntary task switch. Psychological Science, 15(9), 610–615.
Averbeck, B. B. (2015). Theory of Choice in Bandit, Information Sampling and Foraging Tasks. PLoS Computational Biology, 11(3), 1–28. https://doi.org/10.1371/journal.pcbi.1004164
Behrens, T. E. J., Woolrich, M. W., Walton, M. E., & Rushworth, M. F. S. (2007). Learning the value of information in an uncertain world. Nature Neuroscience, 10(9), 1214–1221. https://doi.org/10.1038/nn1954
Beilock, S. L., & Decaro, M. S. (2007). From poor performance to success under stress: Working memory, strategy selection, and mathematical problem solving under pressure. J Exp Psychol Learn Mem Cogn, 33(6), 983–998. https://doi.org/10.1037/0278-7393.33.6.983
Benson-Amram, S., Dantzer, B., Stricker, G., Swanson, E. M., & Holekamp, K. E. (2016). Brain size predicts problem-solving ability in mammalian carnivores. Proceedings of the National Academy of Sciences, 113(9), 2532–2537. https://doi.org/10.1073/pnas.1505913113
Bilalić, M., McLeod, P., & Gobet, F. (2008). Why good thoughts block better ones: The mechanism of the pernicious Einstellung (set) effect. Cognition, 108(3), 652–661.
Bonawitz, E., Shafto, P., Gweon, H., Goodman, N. D., Spelke, E., & Schulz, L. (2011). The double-edged sword of pedagogy: Instruction limits spontaneous exploration and discovery. Cognition, 120(3), 322–330. https://doi.org/10.1016/j.cognition.2010.10.001
Brosnan, S. F., & Hopper, L. M. (2014). Psychological limits on animal innovation. Animal Behaviour, 92(March), 325–332. https://doi.org/10.1016/j.anbehav.2014.02.026
Cash-Padgett, T., & Hayden, B. (2019). Overstaying in patchy foraging can be explained by behavioral variability [Preprint]. Animal Behavior and Cognition. https://doi.org/10.1101/868596
Charnov, E. L. (1976). Optimal foraging, the marginal value theorem. Theoretical Population Biology, 9(2), 129–136. https://doi.org/10.1016/0040-5809(76)90040-X
Chevin, L.-M., & Hoffmann, A. A. (2017). Evolution of phenotypic plasticity in extreme environments. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1723), 20160138. https://doi.org/10.1098/rstb.2016.0138
Chrysikou, E. G., & Weisberg, R. W. (2005). Following the wrong footsteps: Fixation effects of pictorial examples in a design problem-solving task. J Exp Psychol Learn Mem Cogn, 31(5), 1134–1148. https://doi.org/10.1037/0278-7393.31.5.1134
Cohen, J. D., McClure, S. M., & Yu, A. J. (2007). Should I stay or should I go? How the human brain manages the trade-off between exploitation and exploration. Philosophical Transactions of the Royal Society B: Biological Sciences, 362(1481), 933–942. https://doi.org/10.1098/rstb.2007.2098
Collard, M., Kemery, M., & Banks, S. (2005). Causes of Toolkit Variation Among Hunter-Gatherers: A Test of Four Competing Hypotheses. 19.
Colwell, R. K. (1974). Predictability, Constancy, and Contingency of Periodic Phenomena. Ecology, 55(5), 1148–1153. https://doi.org/10.2307/1940366
Crooks, N. M., & McNeil, N. M. (2009). Increased practice with ‘set’ problems hinders performance on the water jar task. Preceedings of the 31st Annual Conference of the Cognitive Science Society, Cognitive, 643–648.
Dart, R. A., & Salmons, A. (1925). Australopithecus africanus: The man-ape of South Africa. A Century of Nature: Twenty-One Discoveries That Changed Science and the World, 10–20.
Davis, S. J., Schapiro, S. J., Lambeth, S. P., Wood, L. A., & Whiten, A. (2019). Behavioral conservatism is linked to complexity of behavior in chimpanzees (<em>Pan troglodytes</em>): Implications for cognition and cumulative culture. Journal of Comparative Psychology, 133(1), 20. https://doi.org/10.1037/com0000123
Deaner, R. O., Barton, R. A., & Van Schaik, C. (2003). Primate brains and life histories: Renewing the connection. Primates Life Histories and Socioecology, 233–265.
deMenocal, P. B. (1995). Plio-pleistocene African climate. Science, 270(5233), 53–59.
Dingemanse, N. J., & Wolf, M. (2010). Recent models for adaptive personality differences: A review. Philos Trans R Soc Lond B Biol Sci, 365(1560), 3947–3958. https://doi.org/10.1098/rstb.2010.0221
Dingemanse, N. J., & Wolf, M. (2013). Between-individual differences in behavioural plasticity within populations: Causes and consequences. Animal Behaviour, 85(5), 1031–1039. https://doi.org/10.1016/j.anbehav.2012.12.032
Doebel, S., & Zelazo, P. D. (2015). A meta-analysis of the Dimensional Change Card Sort: Implications for developmental theories and the measurement of executive function in children. Dev Rev, 38, 241–268. https://doi.org/10.1016/j.dr.2015.09.001
Duckworth, R. A. (2010). Evolution of Personality: Developmental Constraints on Behavioral Flexibility. The Auk, 127(4), 752–758. https://doi.org/10.1525/auk.2010.127.4.752
Duncker, K., & Lees, L. S. (1945). On problem-solving. Psychological Monographs, 58(5), i–113. https://doi.org/10.1037/h0093599
Fawcett, T. W., Hamblin, S., & Giraldeau, L.-A. (2013). Exposing the behavioral gambit: The evolution of learning and decision rules. Behavioral Ecology, 24(1), 2–11. https://doi.org/10.1093/beheco/ars085
Fischhoff, B., & Broomell, S. B. (2020). Judgment and Decision Making. Annual Review of Psychology, 71(1), 331–355. https://doi.org/10.1146/annurev-psych-010419-050747
Floresco, S. B. (2011). Neural circuits underlying behavioral flexibility: Multiple brain regions work together to adapt behavior to a changing environment. Psychological Science Agenda, 25(4), 1–8.
Fogarty, L., Creanza, N., & Feldman, M. W. (2015). Cultural Evolutionary Perspectives on Creativity and Human Innovation. Trends in Ecology & Evolution, 30(12), 736–754. https://doi.org/10.1016/j.tree.2015.10.004
Fox, C. R., & Weber, M. (2002). Ambiguity aversion, comparative ignorance, and decision context. Organizational Behavior and Human Decision Processes, 88(1), 476–498. https://doi.org/10.1006/obhd.2001.2990
Froese, T., & Leavens, D. A. (2014). The direct perception hypothesis: Perceiving the intention of another’s action hinders its precise imitation. Front Psychol, 5, 65–65. https://doi.org/10.3389/fpsyg.2014.00065
German, T. P., & Barrett, H. C. (2005). Functional Fixedness in a Technologically Sparse Culture. American Psychological Society, 16(1), 1–4.
German, T. P., & Defeyter, M. A. (2000). Immunity to functional fixedness in young children. Psychonomic Bulletin & Review, 7(4), 707–712.
Gigerenzer, G., Hertwig, R., & Pachur, T. (Eds.). (2011). Heuristics: The foundations of adaptive behavior. Oxford University Press.
Gittins, J. C., & Jones, D. M. (1979). A dynamic allocation index for the discounted multiarmed bandit problem. Biometrika, 66(3), 561–565. https://doi.org/10.1093/biomet/66.3.561
Gopnik, A, Griffiths, T. L., & Lucas, C. G. (2015). When Younger Learners Can Be Better (or at Least More Open-Minded) Than Older Ones. Current Directions in Psychological Science, 24(2), 87–92. https://doi.org/10.1177/0963721414556653
Gopnik, Alison. (2020). Childhood as a solution to explore–exploit tensions. Philosophical Transactions of the Royal Society B: Biological Sciences, 375(1803), 20190502. https://doi.org/10.1098/rstb.2019.0502
Grove, M. (2011). Speciation, diversity, and Mode 1 technologies: The impact of variability selection. Journal of Human Evolution, 61(3), 306–319. https://doi.org/10.1016/j.jhevol.2011.04.005
Hanus, D., Mendes, N., Tennie, C., & Call, J. (2011). Comparing the performances of apes (Gorilla gorilla, Pan troglodytes, Pongo pygmaeus) and human children (Homo sapiens) in the floating peanut task. PLoS One, 6(6), e19555–e19555. https://doi.org/10.1371/journal.pone.0019555
Henrich, J., & McElreath, R. (2003). The evolution of cultural evolution. Evolutionary Anthropology: Issues, News, and Reviews, 12(3), 123–135. https://doi.org/10.1002/evan.10110
Hill, K., Barton, M., & Hurtado, A. M. (2009). The emergence of human uniqueness: Characters underlying behavioral modernity. Evolutionary Anthropology: Issues, News, and Reviews, 18(5), 187–200. https://doi.org/10.1002/evan.20224
Hoehl, S., Keupp, S., Schleihauf, H., McGuigan, N., Buttelmann, D., & Whiten, A. (2019). ‘Over-imitation’: A review and appraisal of a decade of research. Developmental Review, 51, 90–108. https://doi.org/10.1016/j.dr.2018.12.002
Hopper, L. M., Schapiro, S. J., Lambeth, S. P., & Brosnan, S. F. (2011). Chimpanzees’ socially maintained food preferences indicate both conservatism and conformity. Anim Behav, 81(6), 1195–1202. https://doi.org/10.1016/j.anbehav.2011.03.002
Horner, V., & Whiten, A. (2005). Causal knowledge and imitation/emulation switching in chimpanzees (Pan troglodytes) and children (Homo sapiens). Anim Cogn, 8(3), 164–181. https://doi.org/10.1007/s10071-004-0239-6
Hrubesch, C., Preuschoft, S., & van Schaik, C. (2009). Skill mastery inhibits adoption of observed alternative solutions among chimpanzees (Pan troglodytes). Anim Cogn, 12(2), 209–216. https://doi.org/10.1007/s10071-008-0183-y
Ionescu, T. (2017). The variability-stability-flexibility pattern: A possible key to understanding the flexibility of the human mind. Review of General Psychology, 21(2), 123–131. https://doi.org/10.1037/gpr0000110
Jacobson, S. L., & Hopper, L. M. (2019a). Hardly habitual: Chimpanzees and gorillas show flexibility in their motor responses when presented with a causally-clear task. PeerJ, 7, e6195–e6195. https://doi.org/10.7717/peerj.6195
Jacobson, S. L., & Hopper, L. M. (2019b). Hardly habitual: Chimpanzees and gorillas show flexibility in their motor responses when presented with a causally-clear task. PeerJ, 7, e6195–e6195. https://doi.org/10.7717/peerj.6195
Jansson, D. G., & Smith, S. M. (1991). Design fixation. Design Studies, 12(1), 3–11.
Kahneman, D., Slovic, S. P., Slovic, P., & Tversky, A. (1982). Judgment under uncertainty: Heuristics and biases. Cambridge university press.
Kummer, H., & Goodall, J. (1985). Conditions of innovative behavior in primates. Philos Trans R Soc Lond, 308, 203–214.
Laland, K., Matthews, B., & Feldman, M. W. (2016). An introduction to niche construction theory. Evolutionary Ecology, 30, 191–202. https://doi.org/10.1007/s10682-016-9821-z
Laland, K. N., & Reader, S. M. (1999). Foraging innovation in the guppy. Animal Behaviour, 57, 331–340.
Larcom, S., Rauch, F., & Willems, T. (2017). The benefits of forced experimentation: Striking evidence from the London underground network. The Quarterly Journal of Economics, 132(4), 2019–2055.
Laughlin, S. B., & Sejnowski, T. J. (2003). Communication in Neuronal Networks. Science, 301(5641), 1870–1874. https://doi.org/10.1126/science.1089662
Laureiro-Martínez, D., & Brusoni, S. (2018). Cognitive flexibility and adaptive decision-making: Evidence from a laboratory study of expert decision makers. Strategic Management Journal, 39(4), 1031–1058. https://doi.org/10.1002/smj.2774
Lefebvre, L., Reader, S. M., & Sol, D. (2004). Brains, Innovations and Evolution in Birds and Primates. Brain, Behavior and Evolution, 63(4), 233–246. https://doi.org/10.1159/000076784
Lemaire, P., & Leclère, M. (2014). Strategy repetition in young and older adults: A study in arithmetic. Developmental Psychology, 50(2), 460–468. https://doi.org/10.1037/a0033527
Luchins, A. S. (1942). Mechanization of problem solving: The effect of Einstellung. Psychological Monographs, 54(6), 1–95.
Luchins, A. S., & Luchins, E. H. (1950). New experimental attempts at preventing mechanization in problem solving. The Journal of General Psychology, 42, 279–297.
Mata, R., Wilke, A., & Czienskowski, U. (2013). Foraging across the life span: Is there a reduction in exploration with aging? Frontiers in Neuroscience, 7. https://doi.org/10.3389/fnins.2013.00053
McFadden, D., Machina, M. J., & Baron, J. (1999). Rationality for economists? In Elicitation of preferences (pp. 73–110). Springer.
Meiran, N. (2010). Task Switching: Mechanisms Underlying Rigid vs. Flexible Self-Control. In R. Hassin, K. Ochsner, & Y. Trope (Eds.), Self Control in Society, Mind, and Brain (pp. 202–220). Oxford University Press. https://doi.org/10.1093/acprof:oso/9780195391381.003.0011
Milton, K. (1981). Distribution Patterns of Tropical Plant Foods as an Evolutionary Stimulus to Primate Mental Development. American Anthropologist, 83(3), 534–548. https://doi.org/10.1525/aa.1981.83.3.02a00020
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D. (2000). The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cognitive Psychology, 41(1), 49–100. https://doi.org/10.1006/cogp.1999.0734
Monsell, S. (2003). Task switching. Trends in Cognitive Sciences, 7(3), 134–140. https://doi.org/10.1016/S1364-6613(03)00028-7
Muthukrishna, M. (2015). The Cultural Brain Hypothesis and the transmission and evolution of culture.
Muthukrishna, M., Doebeli, M., Chudek, M., & Henrich, J. (2018). The Cultural Brain Hypothesis: How culture drives brain expansion, sociality, and life history. PLoS Computational Biology, 14(11). https://doi.org/10.1371/journal.pcbi.1006504
Navarro, D. J., Newell, B. R., & Schulze, C. (2016). Learning and choosing in an uncertain world: An investigation of the explore–exploit dilemma in static and dynamic environments. Cognitive Psychology, 85, 43–77. https://doi.org/10.1016/j.cogpsych.2016.01.001
Ntamboudila, F. K. (2020, March 24). The King of the Jungle.
Odling-Smee, F. J. (1988). Niche-constructing phenotypes.
O’Farrell, S., Sanchirico, J. N., Spiegel, O., Depalle, M., Haynie, A. C., Murawski, S. A., Perruso, L., & Strelcheck, A. (2019). Disturbance modifies payoffs in the explore-exploit trade-off. Nature Communications, 10(1), 3363. https://doi.org/10.1038/s41467-019-11106-y
Öllinger, M., Jones, G., & Knoblich, G. (2008). Investigating the effect of mental set on insight problem solving. Experimental Psychology, 55(4), 269–282. https://doi.org/10.1027/1618-3169.55.4.269
Payne, J. W., Bettman, J. R., & Johnson, E. J. (1994). The Adaptive Decision Maker. The Journal of the Operational Research Society, 45(7), 850–850. https://doi.org/10.2307/2584298
Peterson, E., & Verstynen, T. (2019). A way around the exploration-exploitation dilemma. https://doi.org/10.32470/ccn.2019.1365-0
Pope, S. M. (2018). Differences in Cognitive Flexibility Within the Primate Lineage and Across Human Cultures: When Learned Strategies Block Better Alternatives.
Pope, S. M., Fagot, J., Meguerditchian, A., Washburn, D. A., & Hopkins, W. D. (2019). Enhanced Cognitive Flexibility in the Seminomadic Himba. Journal of Cross-Cultural Psychology, 50(1), 47–62. https://doi.org/10.1177/0022022118806581
Pope, S. M., Fagot, J., Meguerditchian, A., Watzek, J., Lew-Levy, S., Autrey, M. M., & Hopkins, W. D. (2020). Optional-switch cognitive flexibility in primates: Chimpanzees’ (Pan troglodytes) intermediate susceptibility to cognitive set. Journal of Comparative Psychology, 134(1), 98–109. https://doi.org/10.1037/com0000194
Pope, S. M., Meguerditchian, A., Hopkins, W. D., & Fagot, J. (2015). Baboons (Papio papio), but not humans, break cognitive set in a visuomotor task. Anim Cogn, 18(6), 1339–1346. https://doi.org/10.1007/s10071-015-0904-y
Potts, R. (1996). Evolution and Climate Variability. Science, 273(5277), 922–923. https://doi.org/10.1126/science.273.5277.922
Potts, R. (2012). Environmental and behavioral evidence pertaining to the evolution of early Homo. Current Anthropology, 53(SUPPL. 6). https://doi.org/10.1086/667704
Potts, R., & Faith, J. T. (2015). Alternating high and low climate variability: The context of natural selection and speciation in Plio-Pleistocene hominin evolution. Journal of Human Evolution, 87, 5–20. https://doi.org/10.1016/j.jhevol.2015.06.014
Price, E. E., Lambeth, S. P., Schapiro, S. J., & Whiten, A. (2009). A potent effect of observational learning on chimpanzee tool construction. Proc Biol Sci, 276(1671), 3377–3383. https://doi.org/10.1098/rspb.2009.0640
Reader, S M, & Laland, K. N. (2001). Primate innovation: Sex, age and social rank differences. International Journal of Primatology, 22(5), 787–805.
Reader, Simon M., & Laland, K. N. (2002). Social intelligence, innovation, and enhanced brain size in primates. Proceedings of the National Academy of Sciences of the United States of America, 99(7), 4436–4441. https://doi.org/10.1073/pnas.062041299
Reader, Simon M., & MacDonald, K. (2003). Environmental Variability and Primate Behavioural Flexibility. In Simon M. Reader & K. N. Laland (Eds.), Animal Innovation (pp. 83–116). Oxford University Press. https://doi.org/10.1093/acprof:oso/9780198526223.003.0004
Riotte-Lambert, L., & Matthiopoulos, J. (2020). Environmental Predictability as a Cause and Consequence of Animal Movement. Trends in Ecology & Evolution, 35(2), 163–174. https://doi.org/10.1016/j.tree.2019.09.009
Sakai, K. (2008). Task set and prefrontal cortex. Annu Rev Neurosci, 31, 219–245. https://doi.org/10.1146/annurev.neuro.31.060407.125642
Schillemans, V. (2011). The Perseveration Effect in Individuals’ Strategy Choices.
Snell-Rood, E. C. (2013). An overview of the evolutionary causes and consequences of behavioural plasticity. Animal Behaviour, 85(5), 1004–1011.
Sol, D., Duncan, R. P., Blackburn, T. M., Cassey, P., & Lefebvre, L. (2005). Big brains, enhanced cognition, and response of birds to novel environments. Proceedings of the National Academy of Sciences, 102(15), 5460–5465. https://doi.org/10.1073/pnas.0408145102
Sol, D., Sayol, F., Ducatez, S., & Lefebvre, L. (2016). The life-history basis of behavioural innovations. Philos Trans R Soc Lond B Biol Sci, 371(1690). https://doi.org/10.1098/rstb.2015.0187
Speekenbrink, M., & Konstantinidis, E. (2015). Uncertainty and exploration in a restless bandit problem. Topics in Cognitive Science, 7(2), 351–367. https://doi.org/10.1111/tops.12145
Stephens, D. W., & Krebs, J. R. (1986). Foraging theory (Vol. 1). Princeton University Press.
Todd, P. M., & Gigerenzer, G. (2007). Environments That Make Us Smart: Ecological Rationality. Current Directions in Psychological Science, 16(3), 167–171. https://doi.org/10.1111/j.1467-8721.2007.00497.x
Tversky, A., & Kahneman, D. (1974). Judgment under Uncertainty: Heuristics and Biases. Science, New Series, 185(4157,), 1124–1131.
Ueltzhöffer, K., Armbruster-Genç, D. J. N., & Fiebach, C. J. (2015). Stochastic Dynamics Underlying Cognitive Stability and Flexibility. PLOS Computational Biology, 11(6), e1004331. https://doi.org/10.1371/journal.pcbi.1004331
van de Pol, M., Jenouvrier, S., Cornelissen, J. H. C., & Visser, M. E. (2017). Behavioural, ecological and evolutionary responses to extreme climatic events: Challenges and directions. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1723), 20160134. https://doi.org/10.1098/rstb.2016.0134
van Leeuwen, E. J. C., & Call, J. (2017). Conservatism and “copy-if-better” in chimpanzees (Pan troglodytes). Animal Cognition, 20(3), 575–579. https://doi.org/10.1007/s10071-016-1061-7
Van Leeuwen, E. J. C., Cronin, K. A., Schütte, S., Call, J., & Haun, D. B. M. (2013). Chimpanzees (Pan troglodytes) flexibly adjust their behaviour in order to maximize payoffs, not to conform to majorities. PLoS ONE, 8(11), 1–10. https://doi.org/10.1371/journal.pone.0080945
van Schaik, C. P. (2013). The costs and benefits of flexibility as an expression of behavioural plasticity: A primate perspective. Philosophical Transactions of the Royal Society B: Biological Sciences, 368(1618), 20120339. https://doi.org/10.1098/rstb.2012.0339
Vicente, K. J., & Wang, J. H. (1998a). An ecological theory of expertise effects in memory recall. Psychological Review, 105(1), 33–57. https://doi.org/10.1037/0033-295X.105.1.33
Vicente, K. J., & Wang, J. H. (1998b). An ecological theory of expertise effects in memory recall. Psychological Review, 105(1), 33–57. https://doi.org/10.1037/0033-295X.105.1.33
Vitousek, P. M., Mooney, H. A., Lubchenco, J., & Melillo, J. M. (1997). Human Domination of Earth’s Ecosystems. 277, 7.
Vrba, E. S. (Ed.). (1995). Paleoclimate and evolution, with emphasis on human origins. Yale University Press.
Watzek, J., Pope, S. M., & Brosnan, S. F. (2019). Capuchin and rhesus monkeys but not humans show cognitive flexibility in an optional-switch task. Scientific Reports, 9(1), 13195–13195. https://doi.org/10.1038/s41598-019-49658-0
Wertheimer, M. (1945). Productive Thinking. Harper.
Wilson, R. C., Geana, A., White, J. M., Ludvig, E. A., & Cohen, J. D. (2017). Humans use directed and random exploration to solve the explore–exploit dilemma. Journal of Experimental Psychology: General, 143(6), 2074. https://doi.org/10.1037/a0038199
Wright, T. F., Eberhard, J. R., Hobson, E. A., Avery, M. L., & Russello, M. A. (2010). Behavioral flexibility and species invasions: The adaptive flexibility hypothesis. Ethology Ecology & Evolution, 22(4), 393–404. https://doi.org/10.1080/03949370.2010.505580
Wyles, J. S., Kunkel, J. G., & Wilson, A. C. (1983). Birds, behavior, and anatomical evolution. Proceedings of the National Academy of Sciences, 80(14), 4394–4397. https://doi.org/10.1073/pnas.80.14.4394
Zelazo, P. D. (2004). The development of conscious control in childhood. Trends in Cognitive Sciences, 8(1), 12–17. https://doi.org/10.1016/j.tics.2003.11.001
[1] Luchins (1942) referred to cognitive set as ‘einstellung.’ Another common term is conservatism, which is the continued use of a strategy without consideration for alternatives (Brosnan & Hopper, 2014; Davis et al., 2019; Hrubesch et al., 2009).
Mathieu Charbonneau 11 November 2020 (14:16)
Timescales in cognitive flexibility
Thank you, Sarah, for your very stimulating draft.
You often mention time as a key constraint in opting for some strategy (staying, switching, searching), yet timescales are not explicitly discussed. Environmental variability and predictability are timescale relative notions; a sudden flood or the breakdown of a metro station can happen over a few days or weeks; the effects of volcanic eruptions can span over several years; overpopulation, migration, and climate change can span over generations; and so on. Analogously, solutions to problems—finding them, implementing them, etc.—can also be set over different timescales. Devising and implementing novel agricultural techniques and strategies can span multiple seasons and years, whereas finding new baits when fishing or hunting can rapidly be tested and implemented.
I am wondering what sorts of differences various timescales have on the phenomena you discuss, and how it may affect your theoretical framework.
(1) Can you clarify what is the timescale of interest for your framework, and why it is bounded (if bounded)? For instance, you mention the Collective Brain hypothesis and cumulative cultural change. Both of these occur *mostly* over intergenerational timescales, whereas you seem more interested in changes within a lifetime.
(2) Are you thus discussing individual psychology, specifically, or can we also use your framework to speak of strategies adopted by groups and/or whole populations? The discussion of flexibility in Week 6 (Sadie, Manon, Josep) seems particularly relevant for this point.
(3) From a cognitive perspective, timescales seem to play at least two roles: how individuals perceive the timescale of the change could affect their choice in strategy (if seen as a temporary fluke, searching may be done more superficially; vs. entering a new ecosystem as a new permanent home can lead to more search) and the range of memory required for dealing with variable unpredictable environments (e.g., the use of elder knowledge to react to unpredictable change that might have occurred a generation ago).
(4) Timescale may also have a role to play on whether one adopts a random vs. directed exploration strategy for invention. With considerable time to find solutions, random exploration may be useful as it could give useful results on the long run that intense, direct exploration may not. This is what I understand the Cultural Niche/Collective Brain hypothesis to push for. I would definitively be interested in reading your thoughts on this last issue in the section you are considering writing
Dan Sperber 22 November 2020 (23:44)
Reputational costs and benefits?
Thanks you Sarah Pope, for outlining the highly relevant wider framework of the costs and benefits of staying, switching, and searching in shaping cognitive flexibility in humans and other species as a function of the predictability of the environment.
As you have shown in your own work, there are cross-species, cross-cultural and age-related differences in ‘cognitive set’ for which this framework suggested several alternative explanations. “It is possible,” you write, “that unfamiliarity with computer games led Himba participants to consider the game less predictable than their American counterparts. However, another intriguing possibility is that if decision-making heuristics are somewhat domain-general, unpredictability in other areas of Himba participants’ lives might have result in a lowered threshold for exploratory search. This possibility aligns with other research which found that younger children exhibited more exploratory flexibility than adolescents and adults.”
I would like to suggest another kind of cost and benefit – reputational costs and benefits – that may, at least marginally, help explain these differences. Humans involved in cooperative ventures stand to gain or lose by adopting one or another strategy not just by the contribution of this strategy to effectiveness in the pursuit of its direct goal but also by the indication it gives others – potential partners in future cooperation – about the individual's dependability. When this reputational dimension is taken into consideration, rigidity may have an extra benefit of signalling predictability. The switching or searching options, on the other hand, may affect reputation favourably if they turn out to have been clearly justified, or unfavourably if they fail, or if they seem to succeed more through luck than through foresight.
Such reputational concerns are arguably non-existent in less mutually attentive and less cooperative species such as baboons or chimpanzees. Both the importance of reputation and how it can be bolstered or compromise varies across human cultures. In experimental situations which participants, especially in “WEIRD” societies, can interpret as exam-like situations where they are likely to be judged, reputational concerns may play a somewhat different but not less important role.
Do you think that such reputational costs and benefits might contribute to the explanation of the differences in rigidity and flexibility that you discuss? And if so, what light would this throw on flexibility and rigidity not just in the use but also in the transmission of techniques?
Adam Boyette 24 November 2020 (12:10)
Culture in shaping costs/benefits to switching
Thanks so much Sarah for this draft. I find the constrained flexibility framework to be very useful in thinking about how environmental features promote exploration (i.e. exploratory flexibility). Somewhat in relation to both Mathieu's and Dan's comments, I am anxious to know how you might integrate culture into this framework? (I recognize this might pertain to the section you haven't written, so I mean the question to be helpful in brainstorming.)
As Mathieu notes, timescale is important to clarify here, but if we consider individual lifetimes, then I would note that, in additional to social considerations such as reputation, like Dan mentions, cultural context can be critical to one's willingness to explore and change strategies. I’m thinking of cultural level variation in forces like conformism, which may act even independently from the natural environment (e.g. Glowacki & Molleman, 2017). One example on my mind is why it is so hard to get people to wear a face mask in the US despite plenty of information available indicating wearing a mask is being better at preventing the spread of Corona virus than not wearing one. In other words, the environment is stable (or at least predictable) but harsh, and even responsive flexibility is inhibited. Love to hear your thoughts.
Glowacki, L., & Molleman, L. (2017). Subsistence styles shape human social learning strategies. Nature Human Behaviour, 1(5), 0098. https://doi.org/10.1038/s41562-017-0098
Adam Boyette 24 November 2020 (16:55)
Culture in shaping costs/benefits to switching: An addendum
Sarah, I'm leaving this addendum after reading Dietrich's draft, which reminded me that you contributed to Lew-Levy et al. (2020). So, a bit red-faced, I'd like to also more specifically ask how you see socialization -- one of the contributors to the "cultural level variation" I referred to in my original question -- fitting into this theoretical space you've outlined?
Lew-Levy, S., Milks, A., Lavi, N., Pope, S. M., & Friesem, D. E. (2020). Where innovations flourish: An ethnographic and archaeological overview of hunter–gatherer learning contexts. Evolutionary Human Sciences, 2, e31. https://doi.org/10.1017/ehs.2020.35
Helena Miton 27 November 2020 (22:14)
Link between exploration and flexibility?
Thanks Sarah for your draft. Most of the questions I wrote down while reading ended up answered by your terminological note. I think there is definitely some ambiguity as to whether flexibility refers to switching between (more rigidly defined) strategies, or to a more 'online' way to adjust behaviors, and this might actually be a useful way to classify the different approaches and accounts offered by all of us participating to the webinar.
Now, I would like to hear a bit more about what are the links between exploratory behavior and flexibility. As far as I understood, exploratory behavior seems to require, almost by definition, flexibility, as it is a choice to explore an unknown strategy. On the other hand, flexibility can be either responsive or exploratory, so it does not seem to be tied as strongly to exploration. Does this mean that exploration is to be considered, overall, as a subset of flexibility?
Sarah Michelle Pope 21 January 2021 (15:59)
CFF and time scales, social context, and more on terminology.
Hi all, thank you so much for your thoughtful comments and questions. I sincerely apologize for my delay in responding!
In response to Mathieu:
Thank you for challenging me to think about this in more detail. You are right, I’ve been slipping between temporal scales without really addressing the implications. I found your fishing lure example quite nicely suited for a thought experiment. If other resources are scarce (i.e., the consequences of not catching a fish are high), CFF would predict that an individual’s propensity to try a new technique would be suppressed. This might be limited to a single fishing excursion, or over an extended period. However, CFF also predicts that – and this is where I think it gets quite interesting – if at any point subsistence needs are not being met or are *predicted to not be met * perhaps based on downward trend in success or another reliable cue, then responsive flexibility might be evoked in order to switch to another strategy. I agree with you, I think the differential implications of CFF in shorter vs longer time periods depend on memory and the ability to predict future returns on these scales. As a side note, I’m planning to revise my definition of responsive flexibility to include strategy switching in response to failure as well as predicted failure.
In response to Mathieu, Dan, Adam:
Yes, I think you’ve all hit the nail on the head. How CFF interacts with socio-cultural factors is where it gets a little messy. On the society-level, the risks of explorative flexibility can be mitigated by acquiring alternative strategies socially; these are lower risk because the predicted returns are readily observed and they are known to work. Thus, even under harsh conditions, CFF would predict that flexible switching to better socially-acquired strategies would still occur. Dan brings up a great point regarding reputational costs, and I think that this could be thought of as an increase in harshness – because the consequences of failure are higher – and would result in reduced explorative flexibility. In the original water-jar study, Luchins (1942, p. 87-88) points out that a subset of inflexible problem-solvers seemed very much aware of the alternative strategy but did not use it for fear of giving the ‘wrong’ response. Another interesting thought would be to think of children’s explorative problem-solving tendencies (Gopnik, 2020; Ionescu, 2017) as a product of loosened reputational costs? I would have to give this some more thought but I think it is intriguing.
This also falls very much into the same category as Adam’s comments regarding cultural norms, especially surrounding conformity. The mask example is great! Yes, I think culturally transmitted biases towards conservatism or liberalism (to borrow Sadie, Manon, Josep’s terminology) are likely to exert culturally-specific influences on the risks and benefits of explorative flexibility. Especially this year, I’ve thought a lot about whether geographic distributions of conservative/liberal mindsets might be explained by differing longitudinal exposures to variability/harshness...but that’s clearly very speculative. Something else I’ve been thinking about is our propensity to switch *away* from socially-acquired vs asocially-acquired strategies, and how this might be culturally mediated. My research group is running a study on cognitive set resulting from socially-acquired vs asocially-acquired strategies and I’m really hoping we can place our findings into a cultural context once fieldwork resumes.
In response to Helena:
Thanks for addressing the terminology question – I completely agree. As for your question about exploration and flexibility, my intuition is to lump directed-exploration with responsive flexibility and random exploration with explorative flexibility. Although, in the CFF explorative flexibility is really meant to encompass both switching from a working strategy to a known or novel alternative. Now I’m regretting my use of ‘explorative flexibility’ as a synonym for elective or optional flexibility. Perhaps I should consider using responsive vs elective flexibility to avoid further confusion. I’ll give this some more thought because you’re right, exploration as the investigation of the unknown, is inherently flexible but it could occur either out of necessity (responsive) or desire (elective/explorative).
Again, sorry for the delay!